22 Energy for Growth
22.1 History
Only in recent times have sciences such as physics and
chemistry become divorced from practial subjects such as raising
animals for us to eat. People were far more sensible in the old
days. Many of the scientists who were interested in agriculture
were also interested in chemistry or physics, and vice versa.
Thus, in the history of this topic, energy for growth, some of the
great names of physics and chemistry were actively thinking about
biological and agricultural phenomena.
Energy is the
capacity of
doing work, while metabolism is
the overall balance of constructive and
destructive changes within living organisms.
- It is difficult to pinpoint
the earliest origins of the concept of metabolism. Carbon was one of the chemical
elements known in the ancient World and the discoveries of the other
major elements in animal respiration were considerably later: hydrogen in 1766
by Henry Cavendish, nitrogen in 1772 by Daniel Rutherford, and oxygen by Joseph
Priestly in 1774.
- In the period from 1779 to 1784, Lavoisier and Laplace in
France were undertaking experiments similar to those undertaken in a
modern‑day
undergraduate course on metabolism.
- A guinea pig was kept in an ice calorimeter
for 10 hours and the metabolic heat generated by the animal was
measured by the
amount of ice that was melted.
- The amounts of heat and carbon dioxide generated were
compared to the equivalent amounts produced
by combustion of known amounts of charcoal.
- In relation to the amount of carbon
dioxide produced, the amount of heat produced metabolically by the
guinea pig
was somewhat more than that produced by combustion of pure carbon.
Despite this
quantitative inbalance, Lavoisier and Laplace concluded correctly that carbonaceous material from the
guinea pig had undergone some type of combustion
involving air drawn into the animal's lungs.
- It was already known oxygen was
picked up by the blood stream from the air in the
lungs. Lavoisier and Laplace had now found the oxygen was
withdrawn from
the blood stream and utilized by the body tissues.
- The extra heat produced by
the guinea pig was, of course, obtained from the formation of water by
oxidation of hydrogen ‑ a fact Cavendish demonstrated when he burnt
hydrogen and obtained water vapour.
- This was confirmed metabolically by the
observation of air exhaled by the guinea pig having a diminished
proportion
of oxygen, but an increased proportion of carbon dioxide and water
vapour.
- At
about this time, similar experiments were undertaken by Crawford in
Britain.
22.2 The respiratory quotient
The RESPIRATORY QUOTIENT
(RQ) is
the volume of carbion dioxide exhaled divided by the volume of oxygen
used. The RQ was
introduced in 1849 by Regnault and Reiset who devised a closed‑circuit
apparatus for the measurement of oxygen consumption and carbon dioxide
production.
Small animals with
a high surface to volume ratio were found to have a greater
rate of respiration per unit of body weight than large animals.
It was
concluded this was due to the greater rate of heat loss in small
animals.
Although
oxygen
consumption is increased after any large meal, the increase is most
dramatic
after a meal with a high protein content. By the
1860s, calorimeters had been developed
for the combined measurement of oxygen consumption, heat production,
and the
elimination of carbon dioxide and water. With this apparatus, it was
possible to show the RQ's of various energy sources.
Carbohydrates =
1.0
Protein
= 0.8
Fat
= 0.7
- Nitrogen in urine is be used to measure the amount of
protein metabolized.
- A bomb calorimeter measures the heat of combustion
of various foods.
- For starches and fats, both methods of calorimetry (live
animal and bomb) give similar
results, but in
the animal calorimeter less heat is obtained from proteins than
in the bomb calorimeter.
- Thus, as well as a major correction for urea
production, the nitrogen to carbon ratios of other nitrogenous wastes
such as
uric acid and creatinine also hasto be taken into account.
- Creatinine is a
metabolic end product of nonenzymatic dehydration of intracellular
creatine,
largely excreted unchanged by the kidneys.
- From values
determined by calorimetry, it was found the different caloric values
of food substances are caused largely by differences in the
amount of
hydrogen that they contain, as shown by the weight of water vapor that
they
yield on combustion .
- The first calorimeter large enough to hold an adult steer
was
constructed at Pennsylvania State College. In modern closed circuit
respiration chambers for small animals, oxygen is supplied internally
from
cylinders of compressed gas, and carbon dioxide and water are absorbed
for the
calculation of respiratory quotients. Studies on larger animals require
an open
circuit system in which oxygen and carbon dioxide levels of ingoing and
outgoing air are determined on a continuing basis and integrated over
time.
- Heat production and muscular
work are essential for the animal to stay alive, and growing tissues
compete
with each other and with these essential services to obtain chemical
energy
from nutrients circulating in the vascular system.
- Lean meat with a water
content of approximately 80% contains approximately 4.7 kJ/g, while the
energy
content of fat is approximately eight times greater .
- Mammalian
basal metabolic rate is approximately 12.5 kJ per kg0.75 per
hour.
Where
are we going? What is the point of all this? We are trying
to explain fat deposition in meat animals. Look at the feed
energy required to deposit this fat in a meat animal. Look at
what happens to this energy when humans eat it. So, fat is a bad
thing, eh? Let's have meat without fat, eh? Well, don't
forget - meat totally without fat is bland and seems dry, so who wants
to eat it? So, fat is sort of bad, and sort of good - both at the same
time - a lovely problem for the bright and tidy minds of keen students
to ponder.
22.3 Feed intake and digestion
- Animals have considerable
control over the amount and the nature of the things they eat. In many
practical and experimental situations this behavior may be a primary
factor in
the regulation of growth.
- There are a number of possible control systems for the
regulation of energy balance by consumption.
- Control may be passive, as in the increased energy cost
of locomotion as an
animal grows heavier.
- Active controlling elements may involve hypothalamic feed‑back
circuits.
- The
medium of communication may be represented by the levels of circulating
factors
such as glucose, amino acids, fatty acids , and steroids (which are partitioned
between aqueous and lipid compartments of the body).
- Within the central nervous
system, opioid peptides such
as beta-endorphin, methionine‑ and
leucine-enkephalin, and dynorphin
are involved in modulating feed intake.
- Ruminants consume a
considerable volume of roughage, and the rumen may be filled before
energy
requirements are satisfied. Normally, however, feeding behavior changes
to
accomodate differences in the energy content of the feed.
- Control signals may be transmitted
by acetate and propionate sensors in, or near the
rumen.
- But there
is also a complex interaction between diet and the growth of the gut
itself. Diets with high levels of complex carbohydrates and inadequate
protein may cause extra growth of the gastrointenstinal tract.
- Behavioural factors also are
involved in the animal's growth responses to long‑term stress - stress
diminishes live weight gains, mainly because of a decrease in fat
deposition.
22.4 Energy distribution models
Although the growth equation
of von Bertalanffy may be
translated into energy terms, the other growth
equations in which deceleration is a function of body weight or of age
present
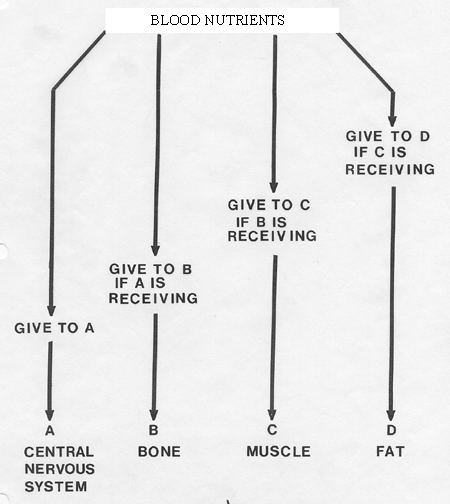
more of a problem. One approach to the problem is given by Sir John
Hammond's
idea of body tissues competing for circulating nutrients.
There
are a number of astute points about this model;
- (1) it concerns itself only
with assimilation,
- (2) it is based on a priority system where vital organs
make
the greatest claim, and
- (3) it encompasses the division of energy between
animal growth and animal reproduction.
In the pregnant female, the uterus attains
a priority for food distribution between A and B. The model also
corresponds fairly well with our present understanding of the endocrine
control
of the flow of energy. In ruminants, for example, the energy flow to
skeletal
muscle is increased by somatotropic hormone (STH) while the flow to adipose
tissue is increased by
insulin . Also, nitric oxide
now is recognised as controlling
blood flow in a number of organs, as in the effect of FGF (fibroblast growth
factor) in modulating blood flow in skeletal muscle . In other
words, circulating
factors may control nutrient availability patterns between
tissues by diverting the flow of blood-borne nutrients, as well as by
switching
on and off the energy assimilation systems in various tissues.
- Insulin
is particularly
important in controlling the distribution of nutrients from the blood
stream
since it turns on the whole energy catching system of a cell.
- At the cell
membrane, insulin increases
glucose and amino acid transport.<>
- In the cytoplasm, insulin activates
ribosomes and mitochondria, while
inhibiting lysosomes .
- In the nucleus, insulin modulates the
synthesis of DNA and RNA.
- Receptors on the cell
membrane recognize insulin molecules and bind them, although some
insulin does
enter the cell, perhaps to exercise its long‑term control functions.
- Insulin
receptors may be heterogeneous in nature with differences occurring in
their
affinity and capacity for binding insulin molecules.
- The concentration of
receptor sites on the cell membrane and the affinity of receptor sites
for
insulin are both variable, and are interrelated with the overall levels
of
insulin in the blood stream.
Hammond's model was also supposed
to explain the effect of reduced energy intake on muscle development.
But this did not survive experimental testing. For example, with
pigs at live weights of 150 kg on a submaintenance
diet, the tissues of the carcass are affected in reverse order
to their order of anatomical development. Fat is reduced most, bone is
reduced least, and muscle is intermediate in reduction. For
another example, with Angus steers on a resricted diet reducing live
weight, bone and connective tissue may be relatively
unaffected, muscle mass may be reduced, but there may be only
relatively slight reductions
in carcass fat content. Chemical analyses shows the loss of muscle
weight is from protein, and not to dehydration or to mobilization of
intramuscular
fat.
- MUSCLE : BONE RATIOS (IMPORTANT
WHEN WE LOOK AT WEIGHTS OF BONE-IN MEAT CUTS).
- Within a breed, muscle to bone
ratios may not be a fixed function of carcass weight, if animals have
been
placed on a low plane of nutrition causing weight loss or
inhibition of
further growth.
- Muscle to bone ratios also may remain
constant despite intensive genetic selection for leaness and growth
rate.
- Dairy and beef breeds may differ in their
muscle to bone ratios as a consequence of selection for milk yield
versus meat
yield.
- Differences in muscle to bone ratios between
breeds of beef cattle appear to be a relic of an earlier period of
breed
improvement when certain breeds were selected for their muscular
strength as
draft animals.
To cope with all these
defects in the original Hammond model, Berg and Butterfield
in 1976
proposed an alternative model of nutrient distribution during growth.
Most researchers still agree with this modification.
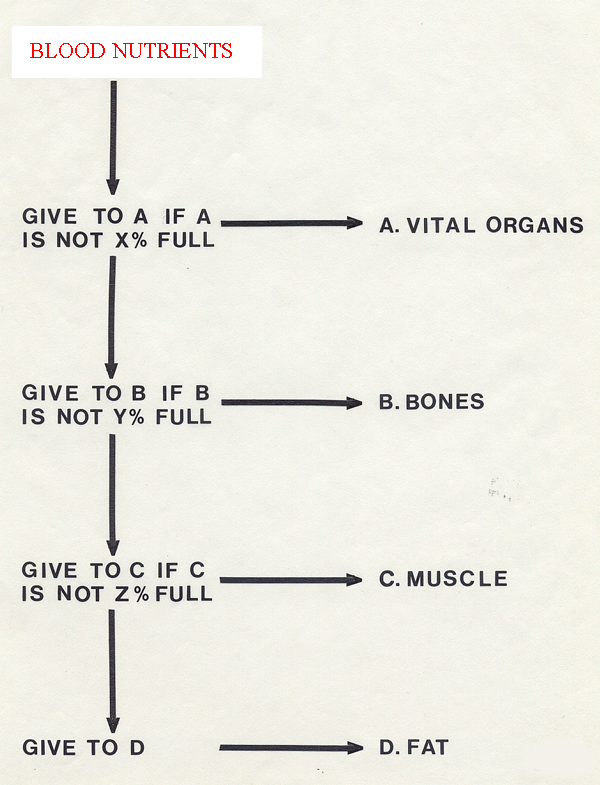
Energy
distribution
between
tissues is now envisaged as a combination of nutrient
availability and tissue
capacity. Hammond's model resembles growth equations in which
specific
growth rate is a function of age, essentially of physiological age. As animals become older they become capable
of reproduction and extensive fat deposition, and their priorities
change
accordingly. Conversely, the model of Berg and Butterfield resembles
growth
equations in which specific growth rate is a function of body size,
because of
the implied limits to tissue volume in all except adipose tissue. Thus,
according to these two models, the
animal "knows" either its
physiological age or what size it should be.
<>This is difficult to
explain! Here are some ideas which have been
proposed.
- TEMPLATE THEORY. One idea is overall body dimensions
might be dependent upon each tissue of
the body regulating its own growth. Each tissue type
produces both templates, with a catalytic action on growth, and
antitemplates.
Templates are thought to be confined to the cell producing them, while
antitemplates diffuse out of the cell to become uniformly diluted
through the
remainder of the body. The concentration of antitemplates in the body
at any
particular time may be a balance between their continuous formation
and degradation. Thus, definitive size is reached when the
concentration of
antitemplates in the whole body finally reaches a level where it blocks
all
intracellular templates.
- TIME
TALLY THEORY. Certain brain cells
may be able to keep track of animal age. These cells might operate a
time tally mechanism matching animal age against the body mass
anticipated
at any particular time. Information on
animal mass may be brought to the time tally by substances resembling
antitemplates produced by one, or a small number of body tissues or
organs. Evidence
from
experimental animals suggests animals do have a "set‑point"
for body weight, and that this may be decreased by experimental lesions
in the
lateral hypothalamus.
22.5 Conclusion
This lecture is more about what we
don't know than what we do know! It might be possible to
combine all these proposed growth‑controlling mechanisms into a single
system
with each tissue compartment being given
- (1) a priority for obtaining energy,
- (2) an essential respiratory requirement, and
- (3) a preferred size calculated
from an interaction between age and body weight.
This could be incorporated
into an algorithm with
- (4) a biological clock,
- (5) an intrinsic system for
monitoring body mass, and logical questions concerning
- (6) pregnancy,
- (7)
required muscular work,
- (8) environmental heat loss,
- (9) nutrient intake and
- (10) stress.
It would, however, be extremely difficult to place
meaningful
limits on parameters 1 to 3, and to program metabolic interactions
resulting
from inputs 4 to 10. The reason is, although the vascular system may
resemble a branching pipeline system, it carries a number of different
fuels at
once. Metabolically, it is compartmentalized (glucose, glycerol, amino acids,
fatty acids, ketone bodies, lactate, pyruvate, high density
lipoproteins, low
density lipoproteins, very low density lipoproteins, etc.) with
a comparable
number of interacting hormonal and biochemical regulators. However, in
cattle,
blood glucose concentration may in fact be correlated with the rate of
growth
in live weight). Required muscular work (7) is a
particularly difficult parameter to evaluate since, as well as
diverting energy
to a mechanical output, work also stimulates muscle growth so that
muscle mass
and physical activity might be correlated.
The real
conclusion? We really do not understand how animals grow, nor how
they distribute nutrients between different types of tissues. We may
think we grow the animals but, really, the animals grow themselves. If
you become an animal scientist, you might help solve some of these
great mysteries!
Further information
Structure and Development of Meat
Animals and Poultry. Pages 463-476.