It is difficult to give a perfect definition of meat animal growth because many of the changes involved are reversible. If an animal increases its live weight by drinking water, do the resulting increments to live weight really constitute growth? Yet, meat often contains 80% water, and it must come from somewhere. If an animal increases its body weight by accumulating fat between and within its muscles, most people might accept that these are true growth increments. Yet, these fat depots readily might be lost if the animal is placed on reduced feed. Likewise, even the myofibrillar proteins of lean meat may be used as an energy reserve in fasting animals, although the growth of the vital organs and nervous system is usually considered to be practically irreversible.
Roughly speaking, growth is an increase in body height, length, girth and weight that occurs when a healthy young meat animal is given adequate food, water and shelter. Live weight is the most important and commonly measured of these features and, if recorded at regular intervals, it may be used to plot a simple growth curve. For the practical purpose of determining feed conversion efficiency, the same data may be expressed as a curve for rate of gain. When live weight data are examined for the small differences in growth (important when large numbers of animals are being raised), even the measurement of live weight becomes a problem, because of weight changes from defaecation, urination and mud on the hide. Thus, in experimental conditions it is preferable to use measurements of body weight mathematically reassembled from the weights of emptied viscera and clean hides obtained after slaughter.
Absolute measurements of body composition can only be
made after an animal has been killed and then, of course, further
growth is impossible. There are several solutions to this problem, but
none of them is entirely satisfactory.
Although studies of carcass composition have an important place in agricultural research, they are not the best means of investigating the process of growth itself. Linear measurements are largely a reflection of skeletal development rather than muscular development, and it is difficult to make accurate skeletal measurements on large animals when they are still alive. With poultry, however, the measurement of linear dimensions may be useful for studying growth and as a basis for genetic selection. Curvilinear measurements made round the girth of the hind-limb of a meat animal are a reasonable index of muscle development, but these are difficult to standardize.
Imagine a simple geometrical structure such as a sphere: if it increases its height, its surface area will increase in proportion to the square of height, while its volume will increase in proportion to the cube of height. If isolated animal cells are grown in suspension in a constantly replenished medium, they form balls of cells. These grow up to several millimetres across, until they reach the limit to which radial diffusion and surface to volume ratio may supply nutrients and oxygen, and the limit at which waste products may still be removed from the center of the ball. If these cells were primitive life forms early in geological time, little further evolutionary increase in size would be possible until they had solved these problems. One solution is gastrulation, to increase the surface to volume ratio by becoming cup-shaped (like pushing a finger into a hollow, soft ball). Another solution is to develop a blood vascular system to enhance the transport of nutrients, oxygen and waste materials. Even for a ball of cells growing in a laboratory dish, however, there are often subtle factors limiting or regulating growth. Biophysical studies with microelectrodes have revealed mechanisms by which cells communicate with each other and "agree" on which cells should divide.
As meat animals grow from birth to slaughter weight, they do not maintain a constant shape. In general, however, even a small increase in linear dimensions causes a proportionately greater increase in body weight because body weight is a function of volume. In newborn farm animals, the rate of heat loss from the body may be a serious problem. In small animals, the surface area for heat loss is relatively high, while the volume of muscle capable of generating heat is relatively small. Since animals maintain an approximately constant body temperature during growth, there must be subtle interactions between body size and the mechanisms which regulate body temperature. Basal metabolism (energy use in a resting animal) is proportional to body weight raised to the power of 0.75, rather than to the power of 0.66, which would be expected on simple geometrical grounds.
Among meat animals, it is difficult to find a situation in which an animal may grow to any marked extent without also exhibiting development of one type or another. Whereas growth may be considered as an increase in height, length, girth or weight, development may be considered as a change in composition, structure or ability, although neither definition is completely satisfactory.
In functional systems such as the locomotory system, development may occur in direct response to growth in live weight. For example, as an animal grows heavier, it may need to use more of its muscle mass to oppose gravity, and there may be changes in the physiological properties of the diverted muscle mass. Similar functional changes occur in the digestive system, bones and other body systems. After growth has ceased in the adult, developmental changes continue as the animal passes through maturity to senescence. Developmental changes are directed towards the attainment of a mature composition, structure or ability. However, the retrogressive changes that occur later in life and which are associated with a decline in composition, structure and ability, are also developmental changes (senescence). Development and senescence are merely the early and late stages of ontogeny, the progress of an individual animal through its life cycle.
Although some aspects of growth such as fat deposition appear
to be reversible, this is rarely true of developmental processes. As an
animal is growing, a vast number of developmental changes are taking
place concurrently. These changes are not usually undone if an animal
simply loses weight. For example, a reversible accumulation of the
triglyceride stored in fat cells may be accompanied by an increase in
the number and size of fat cells, but the loss of a moderate amount of
triglyceride may occur by all cells releasing a share of their
triglyceride. Thus, after "degrowth" appears to have occurred, the
animal has not reverted to the developmental state (adipose cell
number) it had before the period of growth started.
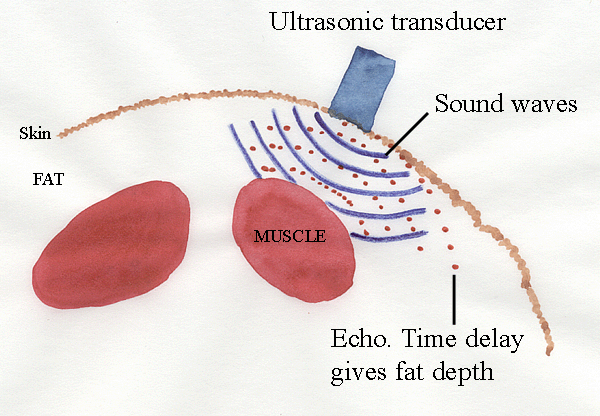
Ultrasonic imaging is widely used for humans, and it is far, far
less
expensive than CAT or NMR. Ultrasonic measurements of fat and muscle
thickness generally are quite reliable and are used routinely in
commercial
agriculture.
The relative development of the carcass also may be studied using specific gravity. Instead of being immersed in water by the classical method of Archimedes, the air displacement technique uses negative air pressures around an animal in an experimental chamber. The accumulation of fat (low density) decreases the overall density of the body so that density then provides an index of fatness. In practice, however, it is difficult to obtain accurate volumetric measurements allowing for the offsetting effects of bones. In cattle, bone density increases with age and varies between sexes and breeds.
An alternative method for adipose tissue volume is based on the fact adipose tissue has a very low water content. Chemicals such as antipyrin or one of its derivatives are injected into the vascular system from whence they diffuse into the total volume of aqueous body fluids. Provided the injected substance is only slowly metabolized or excreted by the body, and provided it becomes adequately dispersed, the extent to which the injected substance becomes diluted indicates the total diluting volume. In general, fat animals have a proportionately smaller aqueous volume than lean animals.
Another indirect method of estimating body composition is based on the fact nearly all the potassium in the body is intracellular. Since adipose cells contain only a trace of potassium in their scanty amount of cytoplasm, the total amount of potassium in the body is approximately proportional to the lean body mass.
The volume of electrolytes in the animal body is estimated from the disturbance that their electrical conductivity creates in an externally applied electromagnetic field. However, this system is adversely affected by any variation of the animal's position and orientation within the electromagnetic field and, consequently, it is not accurate enough for live animals. However, it works very well for boxed beef and carcasses that can be neatly lined up on a conveyer to pass through the machine.
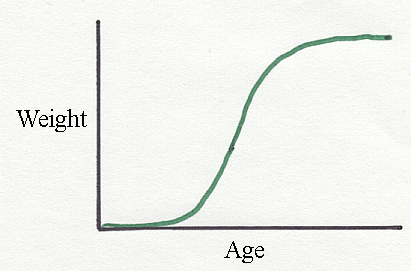
If animal height or weight
are measured from conception to senescence, the
data usually follow a flattened "S" shape called the sigmoid curve.
But the
growth curves of meat animals raised under commercial conditions may
appear as
relatively flat slopes (the middle segment of the flat "S"), and the
sigmoid
shape may only become apparent if the data include very young animals
or
animal beyond a typical market weight. In other words, growth velocity
is
approximately constant during the commercial growing period.
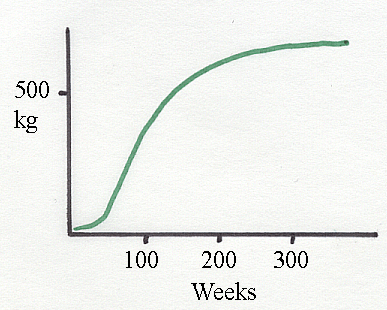
Above is a sigmoid
liveweight growth curve for Holsteins. The "S" shape is far from being exactly "S" shaped.
At first, the fertilized
ovum divides mitotically with little or no growth
in mass. However, as soon as it develops the means of
assimilating energy, the
growth of the embryo accelerates.
At birth and weaning there may be a
temporary deceleration of
growth as an animal switches from one source of
nutrients to another. Except for a slight acceleration at puberty,
subsequent
growth maintains a steady average velocity until the terminal
deceleration
as animals reach their mature size. However, although the average
velocity may be smooth, there may be an underlying circadian (daily)
periodicity in growth rate or even erratic spurts of growth.
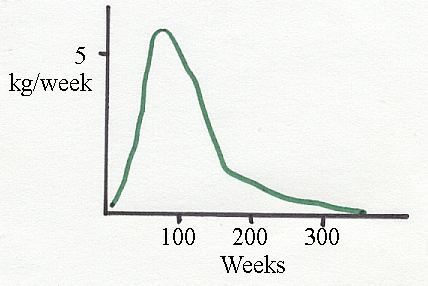
Above is the velocity
curve derived from the sigmoid growth curve for Holsteins.
Anabolism is the building up of body components, while catabolism is their destruction.
Body weight is partitioned between several commercially important compartments. Less energy is required to form a kilogram of lean meat than to form a kilogram of dissectable fat. Thus, a progressive diversion of feed energy away from protein accumulation and towards fat deposition may contribute to the decelerating curve of body weight growth in meat animals, although in consistently fat animals such as pigs this effect may be only slight. The decrease in feed efficiency that occurs with increased live weight in pigs is primarily caused by increased maintenance cost and not increased fat deposition. The time at which the diversion to fat deposition occurs may depend on the breed of animal, and is late in animals with a large adult size. The relative amount of water in the lean tissues of the body decreases with age. The rate of decrease in water content may be altered at weaning and at the onset of sexual maturity.
In attempting to give a biological meaning to the terms b and d in von Bertalanffy's equation, one cannot simply assume the proportional growth of the surface areas of the body keeps pace with the growth of the whole body. In pigs, for example, the small intestine grows rapidly after birth but reaches its mature length between 5 and 6 months of age. A similar early growth attainment of final size is found in the alimentary tract of cattle. Similarly, with heat loss from the body surface, the increased insulation provided by subcutaneous fat must be taken into account. Various regions of the body may differ in their depth of subcutaneous adipose insulation, and there are considerable differences between breeds in the contribution of each body region to the total body area.
There is a resemblance between sigmoid growth curves and those of autocatalytic or self-accelerating chemical reactions in which the product of a simple reaction acts as a catalyst to accelerate further reaction. In a first order chemical reaction, the reaction rate is proportional to only one concentration. For example, in a decomposition reaction the disappearance of the decomposing substance is given by,
Since da/dt, the derivative of a constant is zero,
Robertson's initial chemical reaction catalyzed itself: animals initially add new cells that form further new cells, and for a while the system may behave like a population of microorganisms proliferating in an abundant nutrient medium. Even under these conditions, however, mitotic rates usually show some deceleration. For example, in chick embryos at the 17-somite stage, the number of cells is doubled in 4 hours, but to double the numbers again then takes about 13 hours. As young farm animals grow larger, they consume more feed and store increasing amounts of fat. After a certain point, however, the capacity for energy storage becomes limited by the increasing demands of cellular maintenance and repair, and by the diversion of energy towards reproduction. These self-accelerating and limiting factors work in the same way as those in von Bertalanffy's equation. If the postnatal body tissues of birds and mammals were composed solely of undifferentiated cells like those of the embryo, self-acceleration by cell proliferation might be taken seriously. In meat animals, however, this is not the case.
In the skeletal muscles, massive amounts of protein are assembled into myofibrils. Myofibrils grow in length by forming new sarcomeres at their ends, and they grow radially before they subdivide by longitudinal splitting. New proteins for the myofibril are formed by the activity of ribosomes. Thus, myofibrillar growth per se is not self-accelerating, except in an indirect manner - stronger animals might get more to eat. The growth of adipose tissue is dominated by triglyceride accumulation. Triglyceride accumulation is not self-accelerating, and it provides an even smaller indirect advantage to the animal's growth. Thus, the accelerating phase of growth in meat animals becomes all the more remarkable when we realize that much of the newly added biological material is not self-replicating.
In the equations considered so far, the final deceleration necessary to create a sigmoid curve has originated from, or has been proportional to the enlarged body mass itself. Brody championed another way of thinking about the control of deceleration - to envisage it a function of time, so the growth rate decreases in simple inverse proportion to animal age
Growth curves constructed in this manner, with animal age
as the controlling
factor in deceleration, never
reach a point of zero growth in older animals.
Although perpetual growth may occur in lower vertebrates, until cut
short by
accidental death, there is a finite upper limit to the size of mammals
and
birds. In agriculture, age-based
growth curves may provide a reasonable fit to
growth data since the commercial growing period is in the first half of
the
sigmoid curve.
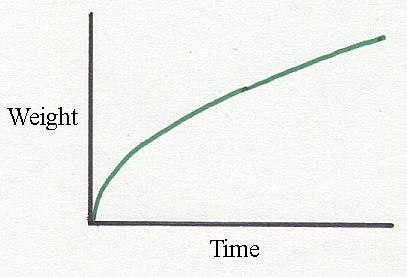
The curve
above shows a typical shape for a time-based growth curve. Note -
it is NOT sigmoid, it never reaches a flat asymptote and
it is not symmetrical.
Instead of growth rate declining in simple inverse proportion to age, it may decline in logarithmic proportion, as in the equation proposed by Gompertz way back in 1825 to calculate financial returns on monetary investments! Thus, in a Gompertz relationship, the logarithm of specific growth rate plotted against time yields a negative linear slope. The Gompertz curve is asymmetric about its inflection point, which occurs early in the curve while the logistic curve is symmetrical.
During the commercial growth of meat animals to market weight, growth may appear almost linear with a constant velocity. During this time, myofibrils, triglyceride and bone matrix accumulate passively by accretionary growth, and the constraints to growth have no measurable impact that may be isolated from sampling error and experimental error. This biologically distinct period of linear growth may be isolated from its sigmoid context to improve the fit of predicted curves to actual data.
In the comparison of breeds differing in their rate of physiological development, it is difficult to justify the use of chronological age rather than physiological age to predict the decline in the logarithm of the specific growth rate. Another problem with time as the regulatory parameter of growth is it makes no allowance for periods when an animal may cease growing because of restricted nutrient intake or stress. This problem does not arise with body weight as the regulatory parameter, and it could also be avoided by using physiological age rather than chronological age. There are, however, no simple units with which to measure physiological age.
The concept of physiological age has been used to create
a physiological
time scale called metabolic age,
based on the relationship between mature size
and the chronological time taken to reach it. The chronological time taken to
reach mature weight is made proportional to the mature weight raised to
the
0.27th power so that the units of metabolic age are those of
chronological age
divided by the 0.27th power of mature weight. Another approach
to the problem
of measuring physiological age is to evaluate animals on the basis of a
set of
criteria changing as animals grow older. Some of the criteria suggested
are:
Genetic selection for leanness in pigs may have delayed their physiological maturation. Thus, genetic selection might be more effective if animals are given a high plane of nutrition and suitable environmental conditions for optimal growth. In model systems, however, the responses to selection appear to be more complex, and genetic advances made under optimal conditions may not automatically be expressed if the animals are switched from optimal to suboptimal environmental growth restraints. One aspect of this problem is physiological maturation may be delayed in a population whose growth is environmentally restrained.
Intrinsic seasonal rhythms in muscle growth are difficult to identify in farm animals reaching market weight in less than a year since intrinsic rhythms are difficult to isolate from seasonal reproductive cycles and seasonal cycles in the nutrient quality of animal feed. In laboratory animal experiments, intrinsic seasonal changes in rates of growth and cellular regeneration have been found to persist even in animals kept under constant conditions of temperature and day length. Seasonal rhythms occur in milk and egg production. In Scottish free-grazing sheep, the average empty body weight of ewes peaks in November and reaches a minimum at the end of April, with subcutaneous fat providing the primary energy reserve depleted during gestation.